У Трансдиференцијација догађа се метаморфоза. Диференциране ћелије датог котиледона трансформишу се у ћелије другог котиледона процесима као што су хистон деацетилација и метилација. Погрешни процеси трансдиференцијације су основа многих болести, као што је Барреттов естрофаг.
Шта је трансдиференцијација?
![]()
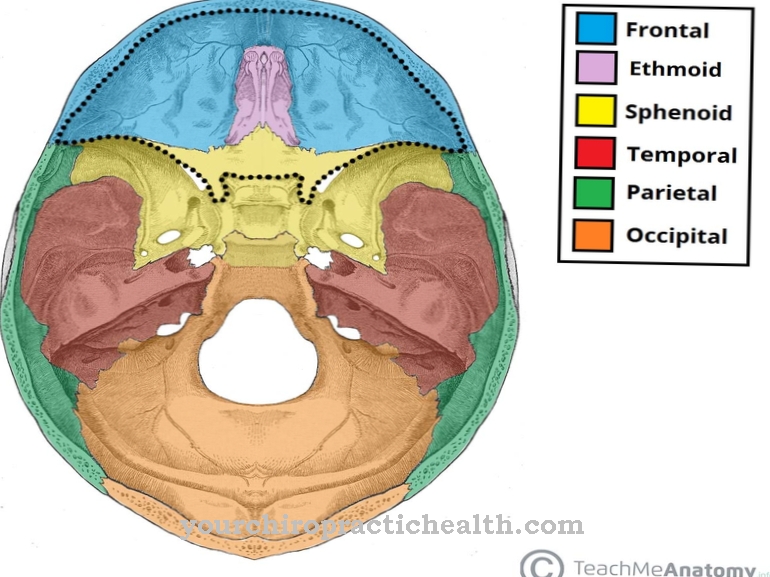
Ембрионални развој одвија се на основу три различита котиледона. Диференцијација је корак у развоју ћелија ембриона. Ћелије се трансформишу у специјализовани облик кроз процесе диференцијације. Прва диференцијација свемоћних ембрионалних ћелија одговара развоју котиледона који су ткивно специфични и стога више нису свемоћни.
Трансдиференцијација је посебан случај или чак реверзија диференцијације. Процес одговара метаморфози. Ћелије једног котиледона се претварају у ћелије другог котиледона. Већина трансдиференцијација се не одвија директно, већ одговара дедиференцијацији, коју заузврат прати диференцијација у супротним смеровима. Научници углавном повезују трансдиференцијацију са људским матичним ћелијама.
Сваком трансдиференцијацијом долази до потпуне промене одговарајуће експресије гена на молекуларно биолошком нивоу. Свака трансдиференцијација захтева промену активности у хиљадама појединих сегмената гена. Процеси патолошке трансдиференцијације одвијају се у вези са неким болестима. У основи, трансдиференцијација не мора имати никакву патолошку вредност.
Функција и задатак
У контексту трансдиференцијације, експресија гена ћелије потпуно се мења на молекуларно генетском нивоу. Ово има импликације на репликацију. У трансдиференцираној ћелији реплицирају се потпуно различити делови гена него што је првобитно замишљено. Из тог разлога, на крају, потпуно другачија синтеза протеина од првобитно планиране.
Трансдиференцијација је праћена деактивацијом претходно активних гена. Ово гашење углавном се одвија путем процеса у контексту хистон деацетилације или метилације на појединим ДНК сегментима. Комплетан процес трансдиференцијације захтева промену активности небројеног броја одсека гена.
Експресија гена трансдиференциране ћелије углавном не одговара оригиналном обрасцу експресије гена у битним деловима. Поступак деацетилације хистона не користи се само за искључивање одређених сегмената гена, већ такође мења способност везања ДНК. Процес деацетилације хистона фокусиран је на хистон, из чије структуре је уклоњена ацетилна група. То даје хистону значајно већи афинитет за ДНК фосфатне групе. Истовремено, постоји мањи капацитет везања између фактора транскрипције и ДНК.
Фактори транскрипције утичу на транскрипцију било позитивно или негативно и били су активатори или супресори. Смањена способност везивања фактора транскрипције резултира инхибицијом експресије појединих гена које се налазе у одговарајућој тачки ДНК.
Процес метилације такође следи принцип инактивације ДНК. Једина разлика је што се процеси метилације фокусирају на метилне групе, а не на хистоне. Ове метилне групе вежу се за одређени део ДНК и на тај начин инактивирају појединачне одсеке ДНК. Када се ћелије диференцирају, њихова експресија гена се значајно мења и многи гени се чак искључују током процеса.
Комплетна трансдиференцијација зависи од високе експресије хиљада гена и истовремено захтева слабу регулацију у експресији на хиљаде других гена. То је једини начин на који су прави протеини на крају доступни да се ћелија трансформише. На пример, мишићној ћелији су потребни битно другачији протеини него ћелија јетре.
Трансдиференцијација се одвија директно или индиректно. Ово скретање одговара дедиференцијацији, након чега слиједи нова диференцијација у другим смјеровима.
Овде можете пронаћи лекове
➔ Лекови за горушицу и надимањеБолести и тегобе
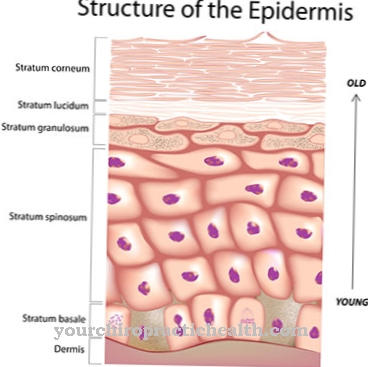
Трансдиференцијације могу бити основа многих различитих болести, што их чини клинички важним. Такозвани Барреттов једњак, на пример, повезан је са процесима трансдиференцијације. Ова болест заснива се на конверзији ћелија епитела, које се током патолошких процеса трансдиференцирају у цревне ћелије које стварају муцин. У овом контексту се говори о цревној метаплазији која је повезана са факултативним ризиком од дегенерације и, на пример, може поспешити развој аденокарцинома. Опћенито, Барреттов синдром описан је као хронична упална промјена дисталног једњака која резултира развојем пептичних чира, што се може појавити као дио компликација код рефлуксне болести. Код синдрома долази до конверзије сквамозног епитела у дисталном једњаку.
Друга болест заснована на трансдиференцираности одговара стварању леукоплакије. Као део ове појаве, оралне ћелије слузнице трансдиферентирају се у преканцерозе, што може промовисати карцином плочастих ћелија. Леукоплакија је хиперкератоза слузокоже која је истовремено истовремено диспластична. Поред усне шупљине, ове леукоплакије јављају се превасходно на уснама и у гениталном пределу. Леукоплакији обично претходи хронична иритација коже или слузокоже. Ова иритација задебљава рожни слој у захваћеном подручју. Црвенкаста слузница постаје толико бјелкаста да се капиларне жиле под густим епителом више не могу разабрати.
Каузални стимулус може бити механичке, биолошке, физичке или хемијске природе. Биолошки подражаји укључују хроничне вирусне инфекције. Хемијски узрочни стимулуси углавном настају пушењем или жвакањем дувана. На пример, слабо пријатна протеза може се сматрати механички узрочно-стимулативним стимулусом.





.jpg)

.jpg)





.jpg)