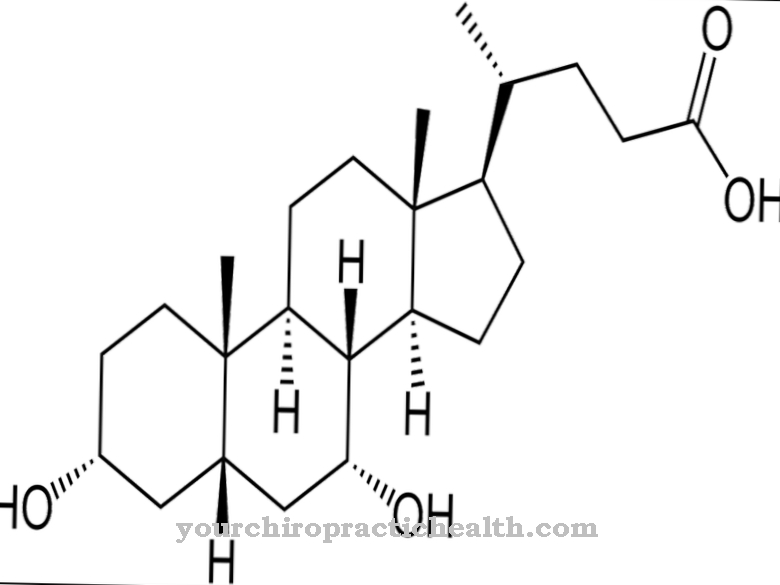
Цитидин припада нуклеозидима и састоји се од нуклеинске базе цитозин и шећерне рибозе. Он формира базни пар са гванозином преко водоничних веза. Такође игра централну улогу у метаболизму пиримидина.
Шта је цитидин?
Цитидин је нуклеозид који се састоји од цитозина и рибозе. Поред аденина, гванина и тимина, цитозин азотне базе је укључен у синтезу нуклеинских киселина. Фосфорилација цитидина ствара цитидин монофосфат (ЦМП), цитидин дифосфат (ЦДП) или цитидин трифосфат (ЦТП).
Цитидин монофосфат је нуклеотид у РНК. Две пуринске и две пиримидинске базе су укључене у структуру нуклеинских киселина, при чему се тимин замењује за урацил у РНК. Аденин и гванин припадају пуринским базама, док тимин, цитозин и урацил припадају пиримидинским базама. Цитидин деаминаза може деаминирати цитидин у уридин. Уридин је нуклеозид произведен од рибозе и урацила. Такође се може фосфорилирати уридин монофосфат.
Уридин монофосфат је такође важан нуклеотид за РНК. Поред тога, ЦДП и ЦТП такође активирају групе за синтезу лецитина, цефлина и кардиолипина. Чисти цитидин присутан је у води растворљивој чврстој супстанци која се распада на 201 до 220 степени. Може се каталитички разградити на цитозин и рибозу помоћу ензима пиримидин нуклеозидаза.
Функција, ефекат и задаци
Цитидин игра централну улогу у метаболизму пиримидина. Пиримидин пружа основну структуру за пиримидинске базе цитозин, тимин и урацил који се јављају у нуклеинским киселинама. Тимин у РНК се замењује за урацил.
Урацил се такође производи деаминацијом цитидина цитидин деаминазом. Хемијске претворбе између три базе пиримидина су од централног значаја за процесе поправљања ДНК и епигенетске промене. У контексту епигенетике, различита својства су модификована утицајем околине. Међутим, генетски материјал се не мења. Модификационе промене организма настају услед различите експресије гена. Процеси диференцијације телесних ћелија за формирање различитих ћелијских линија и органа такође представљају епигенетски процес, у зависности од типа ћелије, различити гени се активирају или деактивирају.
Ово се дешава метилацијом цитидинских база унутар ДНК. За време метилације формира се метилцитозин који се деаминацијом може претворити у тимин. Комплементарни нуклеобаз гванин у супротном двоструком ланцу омогућава препознавање грешке и поновно замену тимина за цитозин. Међутим, гванин се такође може заменити за аденин што доводи до тачкасте мутације. Ако се неметилирани цитозин деаминира, ствара се урацил. Пошто се урацил не појављује у ДНК, одмах га замењује цитозином. Уместо цитозина, стопа мутације услед метилације лагано је повећана.
Истовремено, све више гена се искључује метилацијом тако да ћелије унутар ћелијске линије постају специјализованије. У процесу поправљања, поправни ензими се заснивају на оригиналном ланцу ДНК, који препознају кроз већи степен метилације. Комплементарни низ се такође гради на основу тамо сачуваних информација. Грешке инсталације се исправљају одмах. Поред тога, ензим АИД (Цитиватион-Деаминасе индуцирана активацијом) врло специфично катализује деаминацију цитидинских група у уридинске групе у једноланчаној ДНК. Дешавају се соматске хипермутације, које мењају секвенце антитела из Б ћелија. Тада се бирају одговарајуће Б ћелије. То омогућава флексибилан имунолошки одговор.
Образовање, појава, својства и оптималне вредности
Цитидин је интермедијарни производ метаболизма пиримидина. Као изолована веза, није важно. Као што је већ поменуто, он се састоји од цитозина нуклеинске базе и рибозе шећерне четвороточке. Тело може сам синтетизовати цитозин.
Међутим, његова синтеза је веома енергетски интензивна, тако да се опоравља од градивних нуклеинских киселина у оквиру пута спашавања и може се поново интегрисати у нуклеинске киселине. Када се база потпуно разгради, настају угљендиоксид, вода и уреа. Присутан је као нуклеозид у РНК. У ДНК се цитозин везује за деоксирибозу, тако да се овде нуклеозидни деоксицитидин појављује као грађевни блок.
Болести и поремећаји
Метилације на остацима ДНК из цитидина веома су важне за обележавање ради одвајања различитих биохемијских процеса. Међутим, грешке се могу појавити и код метилације које доводе до болести.
У случају дефектних метилација могу се покренути повећане и смањене активности гена које не задовољавају захтеве. Ови облици метилације преносе се током ћелијске деобе. Дугорочно се дешавају промене које могу довести до болести. На пример, неке ћелије тумора имају различите структуре метилације које се не јављају у здравим ћелијама. На пример, метилација може блокирати одређене гене који кодирају ензиме који регулишу раст. Ако ти ензими недостају, може доћи до не инхибираног раста ћелија. Ово се такође односи на ензиме који иницирају уредну ћелијску смрт (апоптозу) када дође до оштећења ћелија.
Циљано утицај метилације ДНК још увек није могуће. Међутим, постоје студије о потпуној деметилацији туморских ћелија како би се поново подвргле контроли протеина који регулишу раст. Према неколико клиничких студија показало се да деметилација ограничава раст тумора код пацијената са акутном мијелоидном леукемијом. Овај поступак је такође познат и као епигенетска терапија. Процеси метилације такође могу играти улогу код других болести. Због утицаја околине, организам се прилагођава промењеним условима формирањем биолошких модификација заснованих на метилацијама цитидинских остатака ДНК. Тело на тај начин спроводи процес учења, који може, међутим, изазвати и погрешну регулацију.
.jpg)


.jpg)

.jpg)


.jpg)
.jpg)

.jpg)



.jpg)