Сложени ћелијски и физиолошки процеси у живим бићима захтевају фино подешену регулацију на молекуларном нивоу да би се обезбедила прилагодљивост животиње или биљке, на пример, станишту. У ту сврху постоје бројни молекули који интервенишу у процесима као што су ћелијска комуникација, метаболизам или подела ћелије. Један од ових молекула је протеин Цалмодулинкоји уз помоћ калцијума утиче на функцију многих других биолошки активних протеина.
Шта је калмодулин?
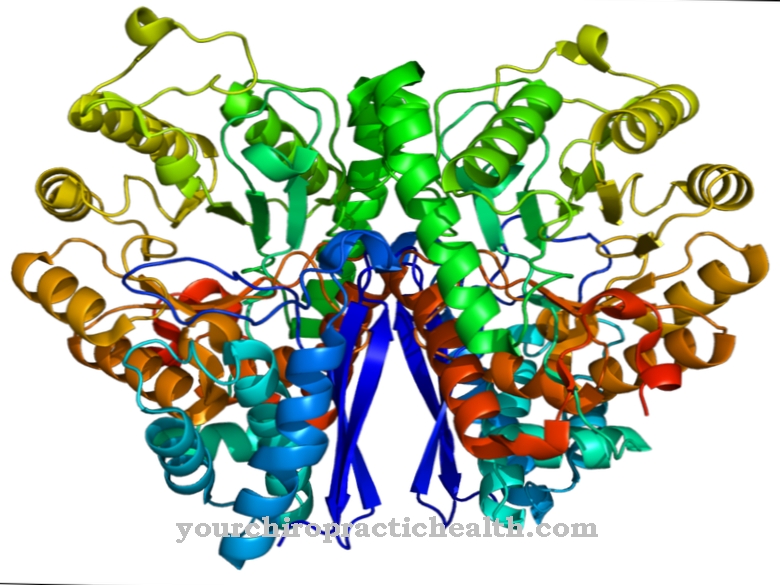
Калмодулин је унутарћелијски регулаторни протеин који везује јоне калцијума. Због своје структуре припада групи протеина ручних ЕФ. Облик калмодулина који се састоји од 148 аминокиселина и дужине је 6,5 нм, подсећа на бучицу. Молекуларна маса овог протеинског молекула је око 17 кДа.
Због своје биолошке функције у преносу сигнала унутар ћелија, калмодулин се такође може класификовати као други гласник, тј. Секундарна супстанцијална супстанца која, међутим, сама по себи није ензиматски активна. У две сферне домене протеина постоје два мотива спирале-хелик на удаљености од 1,1 нм, на које се могу везати укупно четири јона калцијума. Ова структура је позната као ЕФ рука. ЕФ-ручне структуре повезане су водиковим везама између антипаралелних бета листова калмодулина.
Функција, ефекат и задаци
Калмодулину су потребна три до четири везана јона калцијума по молекули да би били активни. У активираном стању, формирани калцијум-калмодулински комплекс укључен је у регулацију великог броја рецептора, ензима и јонских канала са широким спектром функција. Регулисани ензими укључују фосфатазу калцинеурин, који има важну улогу у регулисању имуног одговора, и ендотелну синтезу душичног оксида (еНОС), која производи НО, а који се између осталог користи за опуштање глатких мишића и на тај начин за ширење Крвни судови.
У ниским концентрацијама калцијума активира се и аденилатциклаза (АЦ), а са високим концентрацијама калцијума, с друге стране, ензимски пандан фосфодиестераза (ПДЕ). На овај начин се постиже хронолошки редослед регулаторних механизама: у почетку АЦ креће сигналну путању кроз производњу цикличког АМП (цАМП), касније то поново искључује противнички ПДЕ деградацијом цАМП. Међутим, посебно је познат регулаторни утицај калмодулина на протеинске киназе, као што је ЦаМ киназа ИИ или киоза лаког ланца миозина (МЛЦК), који ће бити детаљније објашњени у даљем тексту.
ЦАМКИИ може да веже остатак фосфата за различите протеине и на тај начин утиче на енергетски метаболизам, пропустљивост за јоне и ослобађање неуротрансмитера из ћелија. ЦАМКИИ се налази у посебно високим концентрацијама у мозгу, где игра важну улогу у пластичности неурона, тј. сви процеси учења. Али калмодулин је такође неопходан за процесе кретања. У мировању је концентрација јона калцијума у мишићној ћелији врло ниска и калмодулин је стога неактиван. Међутим, ако је мишићна ћелија узбуђена, калцијум се улива у ћелијску плазму и, као кофактор, заузима четири места везивања на калмодулину.
Ово сада може активирати киназу лаког ланца миозина која помера контрактилна влакна у ћелији и на тај начин омогућава контракцију мишића. Остали, мање познати ензими који су под утицајем калмодулина су гванилат циклаза, Ца-Мг-АТПаза и фосфолипаза А2.
Образовање, појава, својства и оптималне вредности
Калмодулин се јавља у свим еукариотама, који укључују све биљке, животиње, гљивице и групу амебоидних бића. Будући да је молекул калмодулина у овим организмима обично на релативно сличан начин, може се претпоставити да је то еволуцијски врло стари протеин који је настао у раној фази.
По правилу, калмодулин је присутан у релативно великим количинама у плазми ћелије. На пример, у цитосолу нервних ћелија уобичајена концентрација је око 30-50 уМ, тј. 0,03-0,05 мол / Л Протеин настаје током транскрипције и превођења коришћењем ЦАЛМ гена, од којих су до данас позната три алела, која се називају ЦАЛМ-1, ЦАЛМ-2 и ЦАЛМ-3.
Болести и поремећаји
Постоје неке хемикалије које могу имати инхибиторни ефекат на калмодулин и због тога су познате као инхибитори калмодулина. У већини случајева њихово инхибицијско дејство заснива се на чињеници да они пребацују калцијум из ћелије и на тај начин га уклањају из калмодулина, који је тада присутан само у неактивном стању.
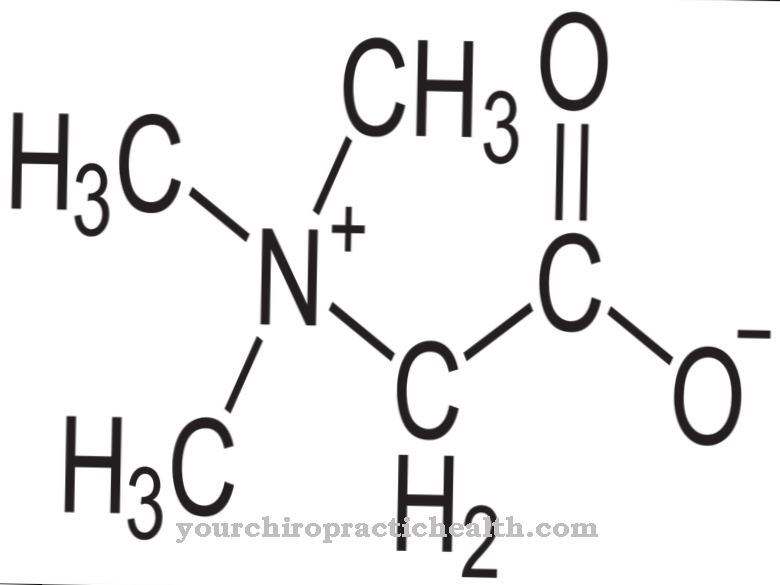
Ове инхибиторне супстанце укључују, на пример, В-7. Поред тога, неки психотропни лекови фенотиазина инхибирају калмодулин. Онолико широке колико су регулаторне функције калмодулина, толико су разнолике замисливе оштећења и поремећаји када протеин не може више да активира кофакторни калцијум, а регулисани циљни ензими су мање активни. На пример, неадекватна активација ЦАМКИИ може довести до ограничења неуронске пластичности, што је основа за процесе учења.
Смањена активација МЛЦК-а смањује контракцију мишића, што може довести до поремећаја покрета. Мања активација ензима калцинеурин услед недостатка калмодулина утицала би на имуни одговор тела, а мање активирање еНО-а водило би нижим концентрацијама НО. Ово последње углавном ствара проблеме где би у супротном душични оксид требао спречити нежељено згрушавање крви и проширити судове у циљу боље циркулације крви. Међутим, у овом тренутку треба такође споменути да сензор калцијума Фрекуенин може под одређеним условима преузети биолошке функције калмодулина и на тај начин заменити молекул.
.jpg)



.jpg)


.jpg)






.jpg)