Тхе ексцитаторни постсинаптички потенцијал је узбудљив потенцијал у постсинаптичкој мембрани неурона. Појединачни потенцијали сумирани су просторно и временски и на тај начин могу створити акциони потенцијал. Поремећаји преноса као што су миастенија гравис или друге миастеније нарушавају ове процесе.
Који је побудни постинаптички потенцијал?

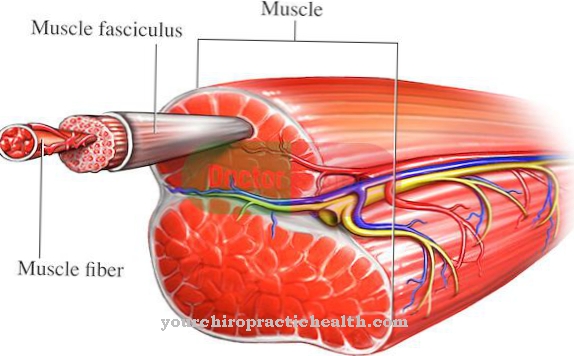
Неурони су одвојени један од другог раздаљином од 20 до 30 нм, познатом и као синаптички јаз. То је минималан јаз између пресинаптичке мембранске регије неурона и постсинаптичке мембранске регије низводне нервне ћелије.
Неурони преносе ексцитацију. Стога је њихов синаптички јаз премоштен ослобађањем биохемијских супстанци, које су такође познате као неуротрансмитери. Ово ствара побудни постинаптички потенцијал на мембранској регији доњеводне ћелије. То је локално ограничена промена потенцијала постсинаптичке мембране. Ова постепена промена потенцијала покреће акциони потенцијал у постсинаптичком елементу. Ексцитаторски постинаптички потенцијал део је спровођења узбуђења неурона и настаје када је деполаризована ћелијска мембрана низводно.
Следећи неурон прима и узбуђује узбудљиве постсинаптичке потенцијале сабирањем и просторно и временски. Када се прекорачи гранични потенцијал ћелије, новоосновани акциони потенцијал однесе се аксоном.
Супротност ексцитацијском постинаптичком потенцијалу је инхибиторни постсинаптички потенцијал. То доводи до хиперполаризације на постсинаптичкој мембрани, што спречава покретање акционог потенцијала.
Функција и задатак
Узбудљив постсинаптички потенцијал и инхибиторни постинаптички потенцијал утичу на све нервне ћелије. Када се премаши њихов праг, нервне ћелије деполаришу. На ову деполаризацију реагују ослобађањем ексцитацијских неуротрансмитера. Одређена количина ових супстанци активира предајно осетљиве јонске канале у неурону. Ови канали су пропусни за јоне калијума и натријума. Локални и градуирани потенцијали у смислу ексцитацијског потенцијала на тај начин деполаризирају постсинаптичку мембрану неурона.
Када се мембрански потенцијал изводи интрацелуларно, ексцитаторски постсинаптички потенцијал је деполаризација сома мембране. Ова деполаризација се дешава као резултат пасивног размножавања. Постоји збир појединачних потенцијала. Количина ослобођеног неуротрансмитера и величина преовлађујућег потенцијала мембране одређују степен ексцитаторног постсинаптичког потенцијала. Што је већа предеполаризација мембране, то је нижи ексцитаторски постинаптички потенцијал.
Ако је мембрана већ деполаризована изнад свог потенцијала за мировање, тада постинаптички побудни потенцијал опада и под одређеним околностима достигне нулу. У овом случају се постиже преокретни потенцијал побудног потенцијала. Ако се предеполаризација покаже још већом, појављује се потенцијал са супротним знаком. Стога се ексцитацијски постинаптички потенцијал не може увијек изједначавати са деполаризацијом. Он се премешта у односу на одређени равнотежни потенцијал, који често остаје испод одговарајућег потенцијала мембране у мировању.
Рад сложеног јонског механизма игра улогу у томе. Уз ексцитаторни постинаптички потенцијал, може се приметити повећана пропусност мембране за калијум и натријум јоне. С друге стране, могу се јавити и потенцијали са смањеном проводљивошћу натријум и калијум јона. Сматра се да је у том контексту механизам јонских канала окидач за затварање свих непропусних калијум јонских канала.
Инхибиторни постсинаптички потенцијал је супротан ексцитаторном постсинаптичком потенцијалу. И овде се мембрански потенцијал локално мења на постсинаптичкој мембрани нервних ћелија. Хиперполаризација ћелијске мембране се дешава на синапси, што инхибира покретање акционих потенцијала у оквиру ексцитаторног постсинаптичког потенцијала. Неуротрансмитери на инхибиторним синапсама покрећу ћелијски одговор. Канали постсинаптичке мембране се отварају и пропуштају калијум или хлоридне јоне. Добивени одлив калијум јона и прилив хлоридних јона изазива локалну хиперполаризацију у постсинаптичкој мембрани.
Овде можете пронаћи лекове
Лекови за слабост мишићаБолести и тегобе
Разне болести нарушавају комуникацију између појединих синапси, а самим тим и трансдукцију сигнала у хемијској синапси. Један пример је неуроткуларна болест миастенија гравис, која утиче на мишићну плочу. То је аутоимуна болест раније непознатог узрока. У случају болести, тело ствара аутоантитела против сопственог ткива. Код мишићних болести, ова антитела су усмерена против постсинаптичке мембране на неуромускуларним крајњим плочама. Најчешћа ауто-антитела код ове болести су антитела за ацетилколински рецептор. Они нападају никотинске ацетилхолинске рецепторе на тачкама везе између живаца и мишића. Настала имунолошка упала уништава локално ткиво.
Као резултат тога, комуникација између нерва и мишића је поремећена, јер интеракција између ацетилхолина и његовог рецептора отежава или чак спречава антитела за ацетилколински рецептор. Акцијски потенцијал због тога више не може прећи из живаца у мишић. Због тога мишић више није узбудљив.
Збир свих ацетилхолинских рецептора смањује се истовремено док се рецептори уништавају имунолошком активношћу. Субсинаптичке мембране се распадају, а ендоцитоза ствара аутофагосом. Транспортне везикуле се спајају са аутофагомсима и ацетилколински рецептори се мењају као резултат ове имуне реакције. Овим изменама мења се и цела плоча на крају мотора. Синаптички јаз се проширио. Из тог разлога, ацетилколин дифузује ван синаптичке пукотине или се хидролизује без везивања за рецептор.
Остале миастеније показују сличне ефекте на синаптичку пукотину и ексцитаторни постсинаптички потенцијал.




.jpg)


.jpg)






.jpg)